FRIDAY, FEB 3, 2023: NOTE TO FILE

St. Matthew Island
Island Earth
Eric Lee, A-SOCIATED PRESS
Abstract: Instead of criticizing a proposed pathway forward by Jack Alpert, as requested, I rewrote it a bit.
COOS BAY (A-P) —
St. Matthew Island: a remote elevation in the Bering Sea, 300 km from the mainland of Alaska.

From: http://ecoglobe.ch/scenarios/e/reindeer.htm
The island of St. Matthew: a distant sight, 300 km from the coast of Alaska.

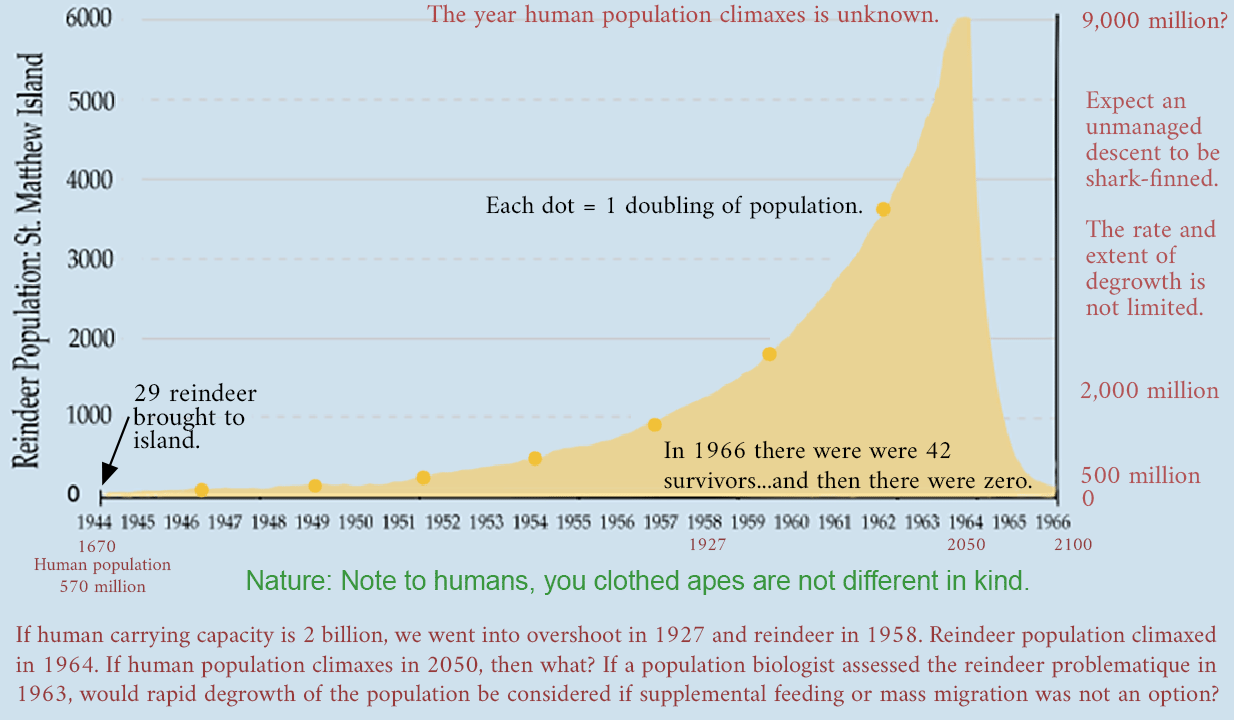
In 1944, the United States Coast Guard established a LORAN station on the island, with 18 personnel (LORAN = long-distance radio navigation system). And 29 reindeer are released on the island as a reserve food source.

In a few months the station is resigned and the men leave the island.

Reindeer find their favorite food — lichens — in abundance on this island.

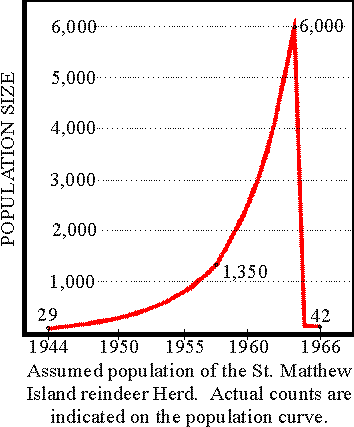
With a surplus to eat and without natural enemies, reindeer reproduce fecundly. 1957: The island is visited by an expedition. In just 13 years, the number of reindeer has increased rapidly from 29 to 1350 animals. The reindeer are in excellent health, fat on their nutritious diet of lichen.

In 1963, scientists revisit St Matthew. In 6 years the reindeer population has exploded from 1350 to 6000 individuals.
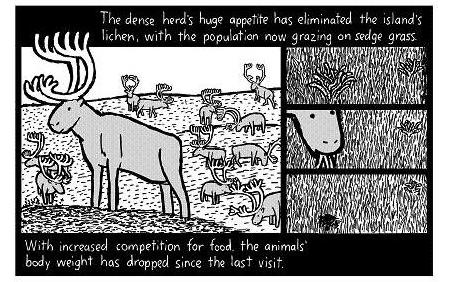
The appetites of the dense herds have eradicated the island’s lichen, and the populace now graze on sedge. With the increased competition for food, the animals have lost weight since the last visit.

In the summer of 1966, the scientists return to the island. They find the island littered with reindeer skeletons. From a peak population of 6,000, only 42 reindeer remain: 41 females and one male. There are no calves.
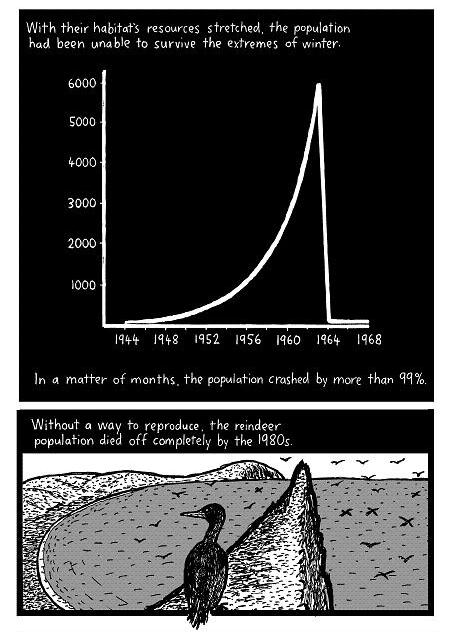
With the tense food situation, the population could not survive the extremes of winter. Without the possibility of producing offspring, the last reindeer died in 1982. In 1987, 22 years after the period of overgrazing, only 10 percent of the original lichen biomass had grown back, and these were mostly lichens that are not preferred by reindeer. It will take many additional “reindeer-free” years before the island will again be capable of supporting a large population of reindeer (potential carrying capacity <900) again.

Untapped natural resources became the reindeer’s source of prosperity — and also the seeds of their death.

The island was only so big and its resources only so much.

Welcome to the Anthropocene.
When where there be too many humans on Earth Island? It will take many centuries before Earth will support a large population of millions of humans again.
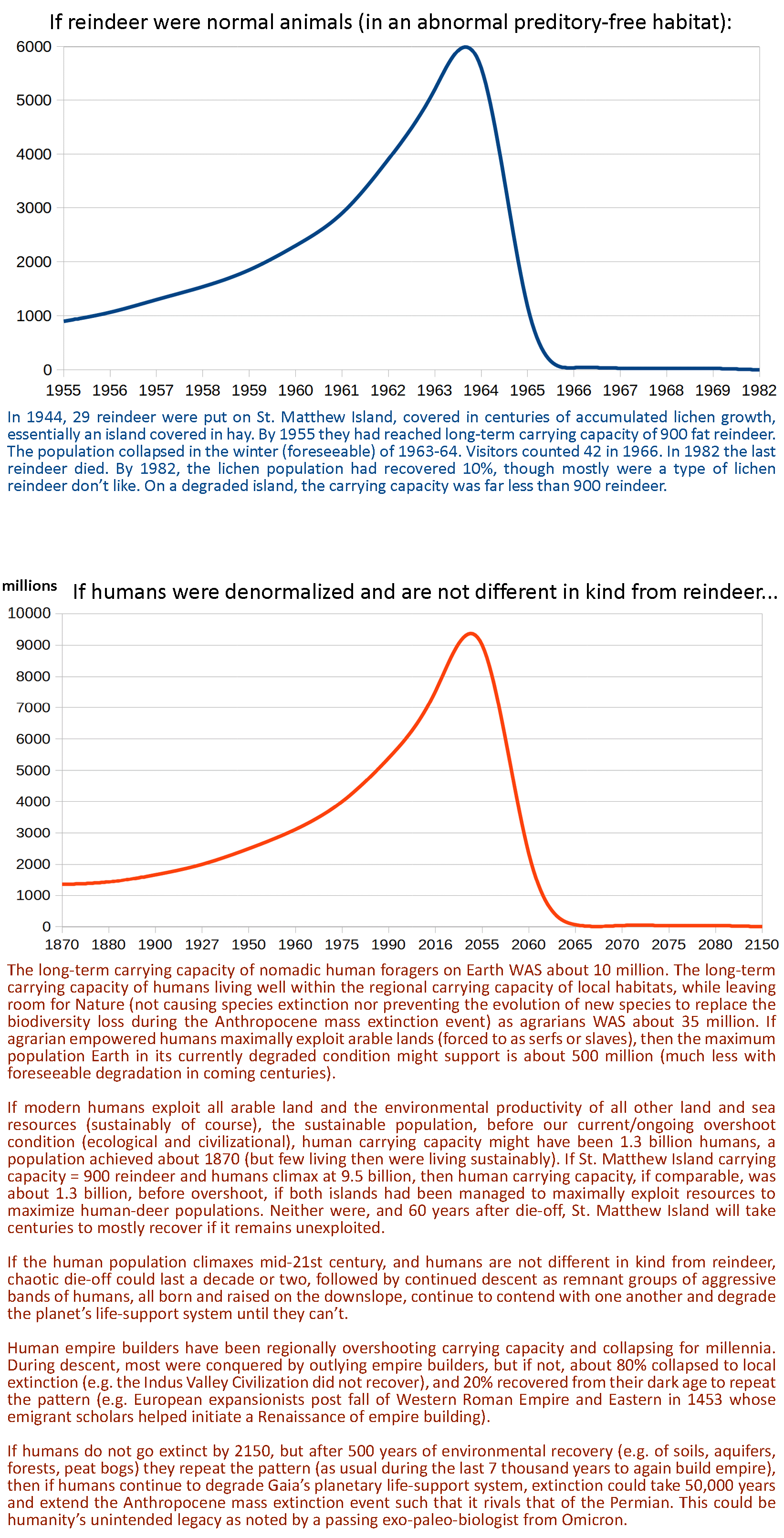

THE INTRODUCTION, INCREASE, AND CRASH OF REINDEER ON ST. MATTHEW ISLAND
By David R. Klein
Alaska Cooperative Wildlife Research Unit, University of Alaska, College
Abstract: Reindeer (Rangifer tarandus), introduced to St. Matthew Island in 1944, increased from 29 animals at that time to 6,000 in the summer of 1963 and underwent a crash die-off the following winter to less than 50 animals. In 1957, the body weight of the reindeer was found to exceed that of reindeer in domestic herds by 24 — 53 percent among females and 46 — 61 percent among males. The population also responded to the high quality and quantity of the forage on the island by increasing rapidly due to a high birth rate and low mortality. By 1963, the density of the reindeer on the island had reached 46.9 per square mile and ratios of fawns and yearlings to adult cows had dropped from 75 and 45 percent respectively, in 1957 to 60 and 26 percent in 1963. Average body weights had decreased from 1957 by 38 percent for adult females and 43 percent for adult males and were comparable to weights of reindeer in domestic herds. Lichens had been completely eliminated as a significant component of the winter diet. Sedges and grasses were expanding into sites previously occupied by lichens. In the late winter of 1963 — 64, in association with extreme snow accumulation, virtually the entire population of 6,000 reindeer died of starvation. With one known exception, all of the surviving reindeer (42 in 1966) were females. The pattern of reindeer population growth and die-off on St. Matthew Island has been observed on other island situations with introduced animals and is believed to be a product of the limited development of ecosystems and the associated deficiency of potential population-regulating factors on islands. Food supply, through its interaction with climatic factors, was the dominant population regulating mechanism for reindeer on St. Matthew Island.
St. Matthew Island, 128 square miles in area and located in the Bering Sea Wildlife Refuge in the north central Bering Sea (Fig. 1), supports a poorly developed land fauna. Native land mammals are restricted to a vole (Microtus abbreviatus) and the arctic fox ( Alopex lagopus ), although a resident population of polar bears (Thalarctos maritimus) existed there in Recent times (Elliot 1882). The reindeer on St. Matthew Island were the result of the release of 24 females and 5 males on August 20, 1944, by the U. S. Coast Guard (Klein 1959). Shortly afterwards, the Coast Guard loran station on the island was abandoned and the island has been uninhabited since then. Specimens taken for study purposes and those shot by Coast Guard personnel as a recreational pursuit have been the only harvest from the herd. With the exception of 10 in 1966, these were all taken during 1957–63 and totaled 105 animals. This paper reports on the population dynamics and range interrelationships of this island reindeer herd from the time of introduction through its rapid increase and crash die-off until July, 1966.
I appreciate the field cooperation of colleagues Dr. Francis H. Fay, Dr. Vernon L. Harms, Jack Manley, and Gerry Cowan and the assistance of James Whisenhant and Dr. Detlef Eisfeld. Dr. Eisfeld also did the analyses of the reindeer ovarian material and, through discussion, provided useful suggestions for the interpretation of the data. Sam Harbo provided advice in the statistical treatment of data.
METHODS
In a preliminary study made during the summer of 1957, a total reindeer count was made, sex and age composition of a sample of the population was determined, specimens were collected, and range studies were conducted. Results of this work have been published (Klein 1959). During the summer of 1963 additional work on St. Matthew Island supplemented the initial study and completed the picture of the course of population growth during the interim 6 years, when population density was high and pressure on the range vegetation was extreme. An attempt was made to visit the island during the summer of 1965, but transportation could not be arranged. In 1966 I again visited the island for 3 weeks in late June and July, during which the surviving reindeer were counted and specimens shot and examined. Skeletal remains from the die-off were identified by sex and age, where possible, and measurements were taken of the antlers and long bones of the hind leg.
Except in 1963, the total counts of reindeer involved a systematic search on foot of the entire island in as short a time as possible to reduce the chance of reindeer movements confusing the counts. Observations usually were made with binoculars and a spotting scope from the ridge tops which divide the island transversely into several broad valleys. Sex and age composition counts were made in conjunction with the total counts and at other times when conditions were favorable for relatively close observation, which was essential for this work. Sex of adults was determined on the basis of the external genitalia and the relative size of the antlers, and fawns, yearlings, and adults were determined on the basis of size and body conformity, although yearlings and adult cows could be differentiated only under the best of circumstances. In 1963, the total count was obtained by completely covering the island with two U. S. Coast Guard helicopters.
Animals taken as specimens were shot from randomly encountered bands, although some selection was involved in attempting to secure representatives of all sex and age groups. Weights were taken with a 200-lb spring scale, which necessitated weighing heavier animals in segments. Standard body measurements were taken and samples of the rumen contents were preserved in formalin for later chemical analysis. The hide, pharyngeal pouches, and nasal cavities were examined for evidence of warble fly larvae ( Oedemagena sp. ) and nasal bots ( Cephenempia sp.), and internal organs were examined for parasites. The entire digestive tract was examined only when the animals were killed close to a supply of water, which was necessary for a thorough examination.
Skeletal remains of reindeer encountered during our work on the island were sexed on the basis of the relative size of the antlers and the character of the pelves. The pelves of adult males had characteristic tuberosities on the posterior border of the ischium and the pubic symphyses were thicker than in the females. Age was determined on the basis of tooth development and wear.
The vegetation of the island was studied through the use of point intercept transects and meter-square quadrats in 1957 and has been reported on earlier ( Klein 1959). Quantitative evaluation of the effects of the reindeer on the vegetation of the island was possible through the use of two groups of meter-square quadrats located in the dry flats, which originally supported the most extensive lichen stands. Each group of quadrats included two which were fenced to protect them from grazing by reindeer and two unprotected plots. Vegetation within the plots was recorded on the basis of the percentage of the total ground cover occupied by each species.
RESULTS AND DISCUSSION Population Dynamics
The original reindeer were obtained from Nunivak Island and were all yearlings; that is, just over 1 year of age. Under the assumed ideal range conditions on St. Matthew Island at the time of the release, the introduced nucleus herd probably increased at a rate approaching the theoretical maximum during the years immediately following. This rate could have been as high as 83 percent the first year if all females bred and produced one fawn each. Under optimum range conditions, reindeer have been known to conceive during their first year and have their first fawns when they are 1 year of age (Palmer 1934, Davey 1963, Skuncke 1967). The actual rates of increase cannot be obtained since data on a yearly basis are not available. Fig. 2 shows the population growth from known counts, from the time of introduction to the summer of 1966.
Age composition counts made in 1963 indicate that the reindeer were experiencing a deceleration in their rate of population growth at that time. In the 1957 counts, the ratio of fawns to adult females was obtained from a sample of 910 animals and the ratio of yearlings to adult females from a sample of 218 reindeer. In these samples there were 75 fawns and 45 yearlings per 100 adult females. In the 1963 counts the ratios were 60 fawns and 26 yearlings per 100 females in samples of 1,652 and 705, respectively. It would be expected that deterioration of the range, with associated reduced physical welfare of the reindeer, would lead to poor fawn survival through their first winter, resulting in low yearling to adult ratios. Calculation of the actual annual rate of increase from these ratios is dependent upon knowledge of the adult sex ratio which was not available in either 1957 or 1963. It was often not possible to distinguish sex and age in the total counts; therefore our composition counts were based on samples of the total population. Because bulls were segregated from females and young when we were on the island, the composition counts did not reflect the true proportion of bulls in the population. However, if one assumes a ratio of 57 bulls:100 cows in the adult segment of the population (the sex ratio of animals over 11/2 years of age among the skeletal material from the die-off ), the net annual increment to the population, after the winter but before the spring fawning period, would have been in the neighborhood of 29 percent in 1957 and 17 percent in 1963.
In 1963 the excessive population on the range was not only reflected in reduced fawn survival, but physical characteristics of the reindeer had changed as well, and the range vegetation had been significantly altered. There was ample evidence of overpopulation and the stage was apparently set for the wholesale die-off that followed. Although no one was present on the island, there is strong presumptive evidence that the dieoff occurred during the winter of 1963 — 64 and apparently largely in the February — April period.
On May 18, 1964, a flight was made over the island in a P2V Navy ice reconnaissance plane in the hope of obtaining a population count of the reindeer through the use of aerial photography. The island was snow-covered with small patches of bare ground visible on wind-swept ridges and south exposures. No reindeer were seen and, even though large groups of reindeer should have been visible under these conditions, we believed that we failed to see them because of the relatively high speed of the plane and reluctance of the pilot to fly at a low altitude over the mountainous terrain. We were not aware, of course, that a die-off had already taken place.
The first realization of the die-off came when Coast Guard personnel went ashore on St. Matthew Island on August 14–16, 1965, to hunt reindeer for recreation. The men hunted in several parties and made a fairly systematic search of the island during three days of unusually favorable weather. On the third day, one group of reindeer, estimated at about 40, was observed near the southeastern end of the island. No other reindeer were seen and the men felt that these were probably the only reindeer remaining on the island. These men also reported that many reindeer skeletons were scattered about, bleached white with no flesh or hide remaining (personal communication ). It seems unlikely that, if the die-off had occurred during the winter of 1964 — 65 rather than 1963 — 64 the skeletons would have been bleached and free of meat and hide to the extent that this report indicated. When we were on the island during 1966, moss had already begun to grow on many of the bones where they were in contact with moist soil, and the skeleton sites, when soil moisture conditions were favorable, were also characterized by lush growth of certain grasses and fortes which have high nitrogen requirements (Fig. 3). These plants are normally restricted to areas adjacent to the bird cliffs which are fertilized by bird droppings, and to disturbed soil near fox burrows and vole colonies. The growth forms of these plants indicated that they had been present for more than one season of growth.
Forty-two live reindeer were on the island when we arrived in 1966. These were counted from the ground in a thorough search of the island and this count (minus 10 animals which were shot) was confirmed from the air on the day we left the island. No fawns or yearlings were present among them and the sample of 10 reindeer also indicates that there had been either no reproduction or no fawn survival since the die-off. Fawns born in the spring of 1964 would have been just over 2 years of age in 1966, and the youngest animal in the sample was just over 4 years of age. The absence of reindeer in the 3-year-old category was also consistent with an assumed 1963–64 die-off since animals in their first winter at the time of the die-off would have been more subject to mortality than prime-age animals.
The winter of 1963–64 on the islands of the Bering Sea was one of the most severe on record from the standpoint of amount and duration of snow on the ground and extreme cold. Weather data were available from the two locations, Mekoryuk on Nunivak Island and St. Paul in the Pribilof Islands, about 250 miles to the east and southeast respectively (U. S. Weather Bureau 1964, 1965). Winter weather conditions on St. Matthew Island are generally milder than on Nunivak which is under a stronger continental influence, and St. Matthew Island has more snow and cold than the Pribilofs which lie almost four degrees of latitude farther south. On Nunivak Island the greatest snow accumulation on the ground during the following months was: January, 31 inches; February, 42 inches; March, 47 inches; and April, 48 inches. This amount of snow for each of these months in 1964 was the greatest during the 20-year record. The average temperature for February was also the coldest on record for that month. At St. Paul Island snow accumulation during the following months was: February, 33 inches; March, 32 inches; and April, 27 inches, also records for this station. Temperatures during February at St. Paul were 7.8 degrees below the monthly mean and were the second coldest in the 20-year record for that month.
With this weather pattern, it is likely that the availability of forage was greatly restricted in late January and February, and the most acute situation was reached sometime after the turn of the year and probably in February or March. That starvation was the cause of death of the reindeer during this period was verified by examination of the medullae of the long bones among the skeletal material. Without exception the cavities were hollow, indicating that no fat was present in the marrow at the time of death. The fat, when present, is resistant to decomposition and will be present in the bones 5 or more years after the death of the animal. This criterion for evaluating physical condition at the time of death has been used successfully among other cervids (Cheatum 1949).
Bones of fetuses were found among the remains of several female reindeer that succumbed during the die-off and they provided a basis for measuring the size of the fetuses at the time of death. Estimated body length of four fetal remains varied from 380 to 480 mm. Davey (1963) obtained body length measurements from 11 fetuses recovered from reindeer shot on St. Paul Island during January and February, 1963, and these varied in size from 8 to 9 inches ( 203–229 mm ) on January 19 to 12 to 15 inches (305 381 mm) on February 15. Since the St. Matthew Island fetuses were only slightly larger than the St. Paul specimens of February 15, it appears that the four adult females carrying the fetuses died in late February and March.
The fact that the reindeer skeletons tended to be grouped ( Fig. 3) suggests that the animals died over a very short period of time; perhaps they were members of a larger band and died during the course of a bedding period. Frequently 10 or more skeletons were found in such close proximity.
The skeletons or other remains of 31 reindeer were found in 1957 and 25 in 1963 and, when possible, they were sexed and aged. No significant differences in the sex and age characteristics were found between the two samples. In both years, animals over 5 years old predominated and males outnumbered females two to one, although some sampling bias undoubtedly existed because the large bleached antlers of bulls are more visible from a distance than are those of cows.
In 1966, skeletal remains of 241 reindeer that had succumbed during the die-off were examined for sex, age, and other characteristics. Only those skeletons from which sex and age could be recorded were included. In the case of animals less than 1 year of age, sex could not be accurately determined. In the overall sample there was a tendency for the small bones of fawns to become scattered and broken and thus less apparent among the tundra vegetation than the skeletons of older reindeer and this probably contributed to a sample bias.
The possibility of a bias among adult animals because of the different methods of sex determination was considered. However, the sex ratio of the subsample identified on the basis of the pelvis was the same as that identified by antlers. Among the sample of 193 animals over 1 year of age which were identified by sex, 41 percent were males and 59 percent females, which is a significant alteration from the assumed equal sex ratio at birth (X2 test; P < 0.02). However, when examined by individual age-classes (Table 1) it is apparent that the sex ratio, essentially equal in the 1 ˝-year class, shifts in favor of females among the 6 1/2 and older animals. This disproportionate adult sex ratio is consistent with the heavier mortality of males indicated by the 1957 and 1963 natural mortality data, and is also consistent with findings among other cervid populations ( Gunvalson et al. 1952 and Klein 1965).
Physical Condition
The physical condition of the reindeer during 1957, 1963, and 1966 showed a pronounced difference. The 1963 animals, in comparable sex and age-classes, were considerably smaller in both body mass and skeletal proportions than the 1957 reindeer. The differences are illustrated in Tables 2 and 3. In Table 3, slopes and elevations of regressions of body characters against age are compared between 1957 and 1963 for the St. Matthew Island reindeer. The significant differences in elevation of regression lines between the two periods indicate that body size differences between the two years are consistent for all age-classes. The differences in slopes, on the other hand, reflect actual differences in growth rates of the reindeer in 1957 and 1963.
In 1957 the St. Matthew Island reindeer were in excellent condition, exceeding weight ranges of reindeer in domestic herds, including average weights from the Nunivak Island herd which was the source of the original animals introduced to St. Matthew Island (Table 4). The large size of the reindeer in 1957 was undoubtedly the result of their recent establishment on a very favorable range (Fig. 4) and the decrease in body weight and skeletal size in 1963 was undoubtedly the product of poorer nutrition, as a result of increased population pressure. However, it is noteworthy that adult weights in 1963, although greatly reduced from 1957, were nonetheless comparable with those from domestic herds, suggesting that in most cases domestic reindeer exist under conditions considerably less than optimal.
Christian (1963) and other workers in the area of physiological mammalogy relate decreasing body size in situations of population increase with density-dependent physiological stress. While there is evidence to support growth depression resulting from stressful situations in laboratory populations of rats and mice (Crew and Mirskaia 1931, Calhoun 1950, Strecker and Emlen 1953), growth of wild ungulates appears to be more directly related to the qualitative and quantitative aspects of the food supply which may not necessarily be related to the density of the population ( Riney 1955, Cowan and Wood 1955, Klein 1964).
The reindeer shot in 1966, all 4 years old or older, represent animals that were alive in 1963 before the die-off. All but one were at least 3 years old in 1963 and therefore had attained essentially all of their growth before the die-off. One would therefore not expect to see any growth response in these animals to the reduced competition for high quality range vegetation, and the body measurements confirm this. The weight of the females in 1966, however, substantially exceeds the average weight for the 1963 adult females. Probably this resulted from reduced competition for high quality forage after the winter of 1963–64 and because the 1966 females had not produced young since the die-off. The absence of the burden of gestation and lactation undoubtedly resulted in a substantial conservation of energy.
The reproductive status of females has been used as an index to physiological welfare ( Cheatum and Severinghaus 1950, Myers and Poole 1962). The reproductive history of female reindeer collected in 1963 and 1966, as determined from ovarian examination, is given in Table S. Unfortunately, ovaries from the 1957 collections were lost in storage. In drawing conclusions from the ovaries, it should be borne in mind that identifying the various ovarian bodies was difficult, owing to variation in their stage of development or retrogression and the lack of suitable reference material; consequently, misinterpretation may have resulted. Also, reconstructing the reproductive history of an animal depends upon accurate aging, and while aging the animals by tooth eruption and wear yielded results consistent with other findings, the method was not based on known-age animals.
Among the females collected in 1963 over 1 year of age, two had corpora albicantia corresponding in number to their age. This may indicate that they conceived as fawns and gave birth to their first young at 1 year of age, although an error may have been made in estimating their age, or one or more of the “corpora albicantia” may have been accessory corpora albicantia of pregnancy similar to those observed among elk ( Halazon and Buechner 1956). Two other females over 1 year of age and the 1-year-old female had apparently not conceived in their first year. The one nonlactating 1963 adult female showed no indication of previous pregnancies although follicular development appeared normal and brown spots, interpreted as scars of erupted follicles, were present throughout the ovaries. There was no apparent explanation for the reproductive failure of this animal as it was in good condition with abundant fat reserves. The fact that two of the 1963 females appeared to have conceived as fawns is surprising. Although breeding of fawns quite likely contributed to the high rate of increase during the early years after the introduction on St. Matthew Island, it seems doubtful that it could have been common during the period of high density when rates of increase were much lower.
The 1966 females apparently produced young before the die-off, with two exceptions: the 4-year-old animal which was a yearling in 1963, and one whose reproductive history is questionable. However, there was no indication that conception took place in any of these animals after the die off. The appearance and number of ovarian brown spots, which were interpreted as scars from erupted follicles, suggest that several ovulations took place without conception occurring during the years following the die-off. It does not appear likely that their failure to breed resulted from nutritional factors. Also physiological stress caused by excess population density, which has been implicated in reproductive failure in rodents and inferred in other mammals (Christian and Davis 1964), should not carry over after the die-off.
Perhaps the most plausible explanation of the lack of reproduction lies among the males rather than the females. In the 1966 sample only one male was collected. No other males were observed among the remaining 32 reindeer although admittedly one or more may have been overlooked owing to small antler size. Normally at this time males would have substantially larger antlers than females, but the antlers of the one male collected were smaller than the average size of the female antlers. It can be speculated, however, that because reproductively active males are more susceptible to winter mortality than females, none of them survived the extreme conditions that brought about the nearly complete annihilation of the herd. If this is true, the absence of pregnancies after the die-off may have been due to the lack of reproductively capable males. The one male known to survive may not have been reproductively active after the die-off. Its relatively low weight and smaller antler growth under conditions which brought about opposite effects in the surviving females tend to support this assumption.
Twelve reindeer specimens in 1957, 15 in 1963, and 10 in 1966 were examined for parasitism. While sample sizes are too small to enable statistical comparisons, lung worms (Dictyocaulus sp.) were found in three of the 1963 animals and in none of the 1957 or 1966 reindeer. None of these infestations were acute. Skin warbles, which commonly infest reindeer and caribou on the mainland, apparently did not establish in the St. Matthew Island herd, for none of these parasites nor the characteristic scars they leave on the skin of the host were found in any of the animals examined.
Lung worm, which has been implicated in mass mortality among other cervids ( Cowan 1951), may have contributed to reduction of the St. Matthew herd, but it certainly was not present in epizootic proportions during the summer preceding the die off Although parasitism may be the actual agent of death in many instances among wild ungulate populations, it is usually associated with debilitation of the animals from other causes such as malnutrition.
The Vegetation
The vegetation on St. Matthew Island is of the arctic tundra type and is of a more xeric nature than that of the Pribilof Islands to the southeast. All plants are low growing and the annual growth of only a few fortes and grasses exceed 30 cm in height. Willows ( Salix spp. ), the only shrubs commonly present, are decumbent in form.
Winter reindeer range on St. Matthew Island is necessarily restricted to windswept areas which are blown free, or nearly free, of snow. Drifted snow collects in stream valleys, depressions, and on the lee side of hills, and greatly restricts availability of vegetation. By nature of their exposure and lack of significant snow cover, the windswept areas support xeric plant communities adapted to the harsh microclimate. A lichen-willow-sedge complex predominated on the winter range through 1957.
Until 1957, the greatest concentration of winter use by reindeer was on the two large dry flats and adjacent low ridges on the southeastern end of the island. Late winter aerial observations made by Rhode in 1955 revealed large numbers of reindeer in these areas ( personal communication ).
Vegetation on the heavily utilized wintering areas adjacent to Big Lake and Cape Upright showed the pronounced effect of heavy reindeer use as early as 1957. Lichen growth had been seriously depleted through the combination of winter grazing, trampling, and shattering, and actual removal of the dry, shattered pieces of lichen by the persistently strong winds. With wind velocities often averaging more than 20 miles an hour during winter months, the potential for plant desiccation and erosion is great. By 1963, lichen growth, which formerly occupied slight depressions between raised hummocks of prostrate willows, had been almost completely eliminated (Fig. 5). In the past, lichen growth apparently was quite similar to ungrazed areas on reindeer-free Hall Island where the lichen mat was 8–12 cm deep (Fig. 6). In 1963 the lichen mat on the old winter range areas seldom exceeded 1 cm in depth and was composed of badly shattered lichens usually unattached to the ground. These same areas showed little change when examined in 1966. The fractured parts of lichens remaining on the ground surface consisted in large part of the nonliving basal parts from which regrowth is not possible. However, there may have been enough living material present to allow for renewal of the lichen mat, but any regrowth will be exceedingly slow. The preferred lichen species, such as Cladonia rangiferina, are the most vulnerable to shattering through trampling, while the more resilient forms which resist shattering, such as Thamnolia vermicularis, are less palatable to reindeer and apparently made up a smaller percentage of the original stands.
Willows fared better than lichens under the heavy reindeer use but by 1963 the willows had begun to show signs of deterioration as a result of heavy winter browsing. It is apparent from observations made in 1957 and on adjacent Hall Island that in the original lichen-willow-sedge stands the willows and sedges were suppressed by the engulfing growth of lichens. The removal of lichens by the reindeer stimulated the growth of willows, sedges, grasses, and some fortes but by 1963 the reindeer were forced to rely heavily on the willows. Comparison of vegetation in fenced and unfenced plots showed an apparent increase in density of willows in the plots that were protected while in the unprotected plots they decreased, but these differences are not statistically significant.
Sedges and grasses continued to increase in density under the continued and increasing grazing pressure. Statistically significant increases in both sedges and grasses were apparent in the fenced and unfenced vegetation plots in 1963 (Tables 6 and 7). Evidence from other reindeer ranges indicates that when lichens are depleted, grasses and sedges are grazed extensively during the winter (Palmer 1929). At the time of the die-off, crowberry (Empetrum nigrum) was apparently the only vegetation available in any volume as evidenced by ruminal material still present among the skeletons.
Summer forage use by reindeer did not result in significant alteration of the vegetation complex. During the 1957 summer field studies, reindeer were observed to make almost exclusive use of the well drained sedge meadows and bog meadows where sedges were very common and were the predominant plants eaten by the reindeer. On the drier, better-drained meadows, Carex nesophila was the most abundant sedge and received the brunt of summer use. The wetter, boggy sites support a wider variety of sedges, but C. aquatilis is usually the dominant form and in these sites it received the heaviest use during 1957. Other sedges, grasses, leaves stripped from willows, and fortes were also important components of the summer diet of the reindeer. All vegetation types received some summer use by reindeer; however, the types with a high proportion of sedges and grasses appeared to support the brunt of summer grazing.
During the 1963 studies, heavy reindeer pressure on the summer forage was evident but no significant lasting damage was noted. Closely cropped sedges and grasses were present in all vegetation types supporting these plants but were most extensive in moist but well-drained meadows, on lake shores and lake floodplains, and on the drier slopes of hills. There appeared to be sufficient summer forage on the island, but competition for the most nutritious and palatable plants was undoubtedly keen. Evidence from other reindeer ranges in Alaska indicates that summer range seldom suffers from overutilization, while winter range condition varies with population pressure and appears to be the most important factor in population control. However, the importance of summer range in the ecology of the reindeer may be greater on an island where opportunity for movement is restricted.
Summer grazing is apparently seldom permanently destructive to moist subarctic tundra. In fact, indications are that limited grazing of the annual growth of sedges, grasses, and some fortes is actually beneficial in stimulating forage production. Harmful effects on summer range are limited to trampling vegetation and compacting loose, moist soil where movements of large numbers of animals are constricted by narrow valleys and other terrain features. Throughout the spring and summer, when actively growing, plants are able to withstand considerable trampling and still recover rapidly. In addition, the high humidity which accompanies the persistent spring and summer fogs on St. Matthew Island keeps the lichens moist and resilient and less subject to shattering than in a drier atmosphere.
The nutritive quality of the low-growing plants which make up the summer forage for reindeer on St. Matthew Island can apparently be very high. This is indicated by the excellent physical status of the reindeer in 1957. On St. Matthew Island the variations in exposure resulting from irregularities in terrain account for a wide range in plant development and maturity. Also, the cool moist summers delay maturity and curing of vegetation. Consequently, during its most nutritious period, the early stages of growth, vegetation is being produced over an extended period. Under the heavy stocking of the range in 1963, competition was apparently great enough to restrict the consumption of highest quality forage by individual reindeer to a minimal portion of their diet.
Selected forage samples collected in 1963 were analyzed chemically for nutritive value and were found to be of relatively high quality. Rumen samples from reindeer shot during 1963 and 1966 were washed to remove the microorganisms and ruminal fluid components and then analyzed chemically as a basis for judging the quality of the forage consumed ( Klein 1962 ). Table 8 shows that for 1963, nutritive values of the washed forage from rumen contents were much lower than the selected forage samples, and apparently reflect the limitation upon the reindeer to select forage qualitatively which was imposed by the high population density. In the 1966 rumen samples, percent protein had increased and percent fiber decreased in comparison with the 1963 samples, apparently as a result of the absence of significant competition among reindeer for the highest quality forage. It is interesting to note in Table 8 that comparative rumen samples from caribou on Adak Island, where these introduced animals are exhibiting remarkable growth rates, show protein and fiber levels even more favorable than those in the 1966 St. Matthew Island samples. In Table 9, comparisons are made between protein and fiber levels of gross rumen samples from St. Matthew Island reindeer and reindeer in a domestic herd on the Seward Peninsula, near Nome. Significantly higher protein levels and lower fiber levels among the St. Matthew Island material, both before and after the die-off, than among rumen samples from the managed reindeer herd emphasize that present domestic reindeer herding practices are seldom based on a knowledge of range ecology.
Range Carrying Capacity
With regard to ungulate range, it can perhaps be stated that forage quantity acts primarily to govern population size while quality determines the size of the individual. Further, in northern regions, food limitation is most critical during the winter period while qualitative variations in the food supply make themselves felt during summer when the physiological demands of animals are highest and growth is most rapid (Klein 1964). Thus range carrying capacity involves two quite different criteria: the winter component which governs the upper limit of the population, and the summer component which determines the physical stature of the individual. Of course, seasonal components of the annual physiological cycle of growth and maintenance in ungulates are not completely independent of one another. Physiological welfare in summer affects reproductive success and winter survival and, hence, rate of population increase; physiological status at the end of winter can influence growth and survival of young and rate and time of initiation of summer fat accumulation in adults.
Various authors have made estimates of grazing capacity for reindeer. Palmer (1929) listed 16–18 reindeer per square mile as the maximum for safe range use and he later (1934) suggested that this might be too high a density for most ranges. Hustich (1951) gave a figure of 13 reindeer to the square mile for the lichen woodland of Labrador. These estimates presumably are for winter range. The density of reindeer on St. Matthew Island in 1957 was 10.5 per square mile and by 1963 it had increased to 46.9 per square mile. It is noteworthy that on St. Paul Island, reindeer reached a density of 49 per square mile just before the “crash die-off’ there in the 1940’s ( Scheffer 1951).
Estimates of desirable stocking on the summer range are generally lacking in the Russian, Scandinavian, and North American literature. Palmer ( 1934: 23–24 ) listed 6 acres (107 reindeer per square mile ) as the combined spring and summer range requirement of reindeer on the basis of studies of fenced reindeer but suggested lower stocking levels on open range. This is not directly comparable to the St. Matthew Island densities because utilizable range is much less than the total land area on the island. Probably as much as half of the island is covered by rock scree which is completely unvegetated except for widely scattered crustose lichens. In addition much of the area utilized as winter range was not used by reindeer in summer.
Mechanisms of Population Control
The reindeer population on St. Matthew Island increased rapidly to a peak which was followed by a crash. This pattern has been observed among other animal populations under varying conditions, but most often among introduced species on islands. Population “explosions” and ensuing die-offs on islands have been reported for reindeer on the Pribilof Islands (Scheffer 1951), cottontail rabbits (Sylvilagus floridanus) on Fishers Island, New York (Smith and Cheatum 1944), and moose ( Alces alces ) on Isle Royale in Lake Superior following their arrival there (Mech 1966). Explanations for this type of response of introduced animals may lie in the characteristics of the new environment or in the lack of plasticity of the introduced species. It is conceivable that an environment containing several diverse potential population-regulating mechanisms (for example, food supply, predators, interspecific competition ) would bring about a more gradual control of an introduced species than an environment with only one limiting mechanism such as food supply. Therefore, one would expect that the more complex the environment in terms of the flora and fauna present, the more graded would be the response of the introduced species. This explanation has been used, of course, to account for the prevalence of cyclic species in northern regions where ecosystems are characteristically less complex than in temperate or tropical regions. Island ecosystems also tend to be less complex than continental ones. Environmental limitations on islands, especially small ones, are finite. There is no continuum from favorable to less favorable habitat for a species occupying an entire island. Island ecosystems, although sparse from a species standpoint owing to restricted access, tend to be younger than continental ecosystems, with the result that there has been less time for the development of complex interrelationships.
Certain species appear less inclined to wide population fluctuations than others when introduced to new environments. The wolves of Isle Royale are a classic example of a new and successful species stabilizing at a level commensurate with the food supply (Mech 1966). There are innumerable instances of the Norway rat, the house mouse, and other species associated with man, gaining access to islands in temperate and tropical regions and, once established, maintaining relatively stable populations. Reindeer have also been introduced to islands without experiencing the wide fluctuations that occurred on St. Matthew Island and the Pribilofs. On Atka and Umnak Islands in the Aleutians, introduced reindeer appear not to have undergone large scale die-offs although accurate records of their population levels are not available. In the Southern Hemisphere the introduced reindeer of South Georgia have apparently stabilized at about 4,000 (Bonner 1958). However, winter climatic conditions are not as severe in the Aleutians or on South Georgia as on St. Matthew Island or the Pribilofs.
There appears to be a relationship between the self-regulatory ability of animal populations and the relative stability of the environments within which they have evolved. For example, the North American deer that are adapted to early successional stages of vegetation, which are of a transitory nature, appear not to have well developed self-regulatory mechanisms and are characterized by wide population fluctuations. On the other hand the roe deer (Capreolus capreolus) in Europe (Andersen 1963) and some bovids, such as the Uganda kob (Adenota kob thomasi) (Beuchner 1963 ), that are found on relatively stable vegetation types, appear to have evolved behavioral mechanisms that tend to contribute to the stability of their populations. The caribou appear to be intermediate in this respect. The tundra and open subarctic lichen forests, which are their native habitat, are climax vegetation types of a very stable nature. Mobility and the development of the migratory habit appear to be compensating mechanisms which prevent destruction of the food resource. In addition, the wolf is an effective predator on caribou and the two species have evolved a relationship that appears mutually beneficial at the level of the population.
On St. Matthew Island and the Pnbilof Islands ( Scheffer 1951), reindeer were introduced to restricted ranges free of predators and subjected to insignificant annual harvests. The normal migratory habit of the species could offer no relief to the pressure of the animals on the range because of the restricted sizes of the islands. The populations expanded rapidly under the good range conditions and winter mortality at first was light because of the abundant lichen forage. Increasing at geometric rates, the populations passed from moderate levels, with respect to the food supply, to excessive populations in only a few years. On St. Matthew Island, and possibly on the Pribilofs as well, the rapid increase to the peak population was coincident with favorable winter climatic conditions.
The large scale die-off of reindeer on St. Matthew Island during the winter of 1963 — 64 was apparently the result of a combination of the following factors: ( 1 ) overgrazing of lichens on the island, which are normally the most important winter forage, by the large numbers of reindeer; (2) excessive numbers of reindeer during the winter of the die-off competing for the very limited available forage; ( 3 ) the relatively poor condition of the reindeer going into the winter as a result of competition for high quality summer forage during the summer of 1963; and (4) extreme weather conditions, particularly deep snow accumulation, during the winter of 1963 — 64, further restricting the availability of the already depleted winter forage. These same factors, jointly operative, were apparently responsible for the reindeer die-off on the Pribilof Islands in the 1940’s ( Scheffer 1951). That weather conditions were not the sole factor in the die-off of reindeer on St. Matthew Island is demonstrated by the good survival of the reintroduced reindeer on St. Paul Island during the same period (F. Wilke in correspondence 1966). Also, on Nunivak Island, although considerable mortality occurred among the reindeer there during the winter of 1983–84, no large die off resulted.
Food supply then, through interaction with climatic factors, was the dominant population-regulating mechanism for reindeer on St. Matthew Island. Other factors of population control, such as disease or parasites and predation, can be ruled out and there is insufficient evidence to suggest that self-regulatory mechanisms of a behavioral (Wynne-Edwards 1965), a genetic ( Chitty 1960 ), or a behavioral-physiological nature (Christian and Davis 1964) were involved in the dieoff.
LITERATURE CITED
ANDERSEN, J. 1963. Populations of hare and roe deer in Denmark. Proc. 16th Internatl. Congr. Zool. 3:347–351.
BONNER, W. N. 1958. The introduced reindeer of South Georgia. Falkland Islands Dependencies Survey Sci. Rept. 22. 11pp.
BUECHNER, H. K. 1963. Territoriality as a behavioral adaptation to environment in Uganda kob. Proc. 16th Internatl. Congr. Zool. 3:5963.
CALHOUN, J. B. 1950. The study of wild animals under controlled conditions. Annals N. W. Acad. Sci. 51: 1113–1112a.
CHEATUM, E. L. 1949. Bone marrow as an index of malnutrition in deer. New York State Conservationist 3(5):19–22.
, AND C. W. SEVERINCHAUS. 1950. Variations in fertility of white-tailed deer related to range conditions. Trans. N. Am. Wildl. Conf. 15:171–190.
CHITTY, D. 1960. Population processes in the vole and their relevance to general theory. Canadian J. Zool. 38(1):99–113.
CHRISTIAN, J. J. 1963. Endocrine adaptive mechanisms and the physiologic regulation of population growth. Pp. 189–353. In W. V. Mayer and R. G. van Gelder (Editors), Physiological mammalogy. Vol. I. Academic Press New York. 381pp.
, AND D. E. DAVIS. 1964. Endocrines, behavior, and population. Science 146(3651): 1550–156.
COWAN, I. McT. 1951. The diseases and parasites of big game mammals of western Canada. Proc. Annul British Columbia Game Convention 5: 37–64.
, AND A. J. WOOD. 1955. The growth rate of the blacktailed deer ( Odocoileus hemionus columbianus ). J. Wildl. Mgmt. 19(3) :331336.
CREW, F. A. E., AND L. MIRSKAIA 1931. The effects of density on an adult mouse population. Biol. Generalis 7:239–250.
DAVEY, S. P. 1963. Reindeer and their management on St. Paul Island, Alaska. Unpub. report, U. S. Burl Commercial Fisheries. Seattle, Washington. 34pp.
ELLIOT H. W. 1882. Report on the seal islands of Alaska. U. S. Commercial Fish and Fisheries Spec. Bull. 176. 176pp.
GUNVALSON, V. E., A. B. ERICKSON, AND D. W. BURCALOW. 1952. Hunting season statistics as an index to range conditions and deer population fluctuations in Minnesota. J. Wildl. Mgmt. 16(2) :121–131.
HALAZON, G. C., AND H. K. BUECHNER. 1956. Postconception ovulation in elk. Trans. N. Am. Wildl. Conf. 21:545–554.
HUSTICH, I. 1951. The lichen woodlands in Labrador and their importance as winter pastures for domesticated reindeer. Acta Geog. 12(1): 1–48.
KLEIN, D. R. 1959. Saint Matthew Island reindeer-range study. U. S. Fish and Wildl. Serv. Spec. Sci. Rept.: Wildl. 43. 48pp.
. 1962. Rumen contents analysis as an index to range quality. Trans. N. Am. Wildl. and Nat. Resources Conf. 27:150–164.
. 1964. Rangerelated differences in growth of deer reflected in skeletal ratios. J. Mammal. 45(2):226–235.
1965. Ecology of deer range in Alaska. Ecol. Monogr. 35(3):259284.
KRAMER, C. Y. 1956. Extension of multiple range tests to group means with unequal numbers of replications. Biometrics 12 (3): 307310.
KREBS, C. J., AND I. McT. COWAN. 1962. Growth studies of reindeer fawns. Canadian J. Zool. 40(5) :863–869.
MECH, L. D. 1966. The wolves of Isle Royale. Fauna of the Natl. Parks of the U. S., Fauna Series 7. 210pp.
MYERS, K., AND W. E. POOLE. 1962. A study of the biology of the wild rabbit, Oryctolagus cuniculus ( L. ) in confined populations. III. Reproduction. Australian J. Zool. 10(2) :225267.
PALMER, L. J. 1929. Improved reindeer handling. U. S. Dept. Agr. Circ. 89. 18pp.
1934. Raising reindeer in Alaska. U. S. Dept. Agr. Misc. Publ. 207. 41pp.
RINEY, T. 1955. Evaluating condition of freeranging red deer (Cercus elaphus), with special reference to New Zealand. New Zealand J. Sci. and Technol. 36(5):429 463.
SCHEFFER, V. B. 1951. The rise and fall of a reindeer herd. Sci. Monthly 73 (6): 356–362.
SKUNCKE, F. 1968. Reindeer ecology and management in Sweden. (In press.) Biol. Papers Univ. Alaska.
SMITH, R. H., AND E. L. CHEATUM. 1944. Role of ticks in decline of an insular cottontail population. J. Wildl. Mgmt. 8(4):311–317.
SNEDECOR, G. W. 1956. Statistical methods. 5th ed. Iowa State University Press, Ames. 534pp.
STEEL, R. G. D., AND J. H. TORRIE. 1960. Principles and procedures of statistics. McGraw-Hill Book Co., Inc., New York. 481pp.
STRECKER, R. L., AND J. T. EMLEN, JR. 1953. Regulatory mechanisms in house-mouse populations: the effect of limited food supply on a confined population. Ecology 34(2):375–385.
U. S. WEATHER BUREAU. 1964. Climatological data Alaska. Vol. 49 (1–13): 1–228.
1965. Climatological data Alaska. Vol. 50(1–13): 1–244.
WYNNE-EDWARDS, V. C. 1965. Self-regulating systems in populations of animals. Science 147(3665): 1543–1548.
Originally published at https://jayhansonsdieoff.net.